

Boreal Forest
Overview
Boreal forest is a conifer or conifer-hardwood forest type occurring on moist to dry sites characterized by species dominant in the Canadian boreal forest. It typically occupies upland sites along shores of the Great Lakes, on islands in the Great Lakes, and locally inland. The community occurs north of the climatic tension zone primarily on sand dunes, glacial lakeplains, and thin soil over bedrock or cobble. Soils of sand and sandy loam are typically moderately acid to neutral, but heavier soils and more acid conditions are common. Proximity to the Great Lakes results in high levels of windthrow and climatic conditions characterized by low summer temperatures and high levels of humidity, snowfall, and summer fog and mist. Additional important forms of natural disturbance include fire and insect epidemics.

Rank
Global Rank: GU - Unrankable
State Rank: S3 - Vulnerable
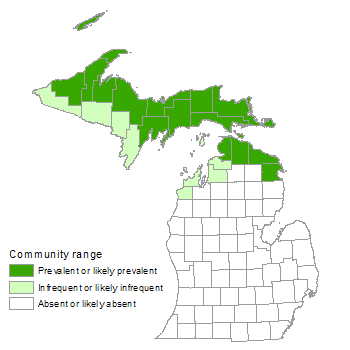
Landscape Context
Boreal forest typically occupies upland sites, often containing local wet depressions, along shores of the Great Lakes, on islands in the Great Lakes (e.g., Isle Royale, Drummond Island, and Beaver Island), and locally inland (e.g., areas within the Negaunee-Michigamme Highlands of the western Upper Peninsula). Coastal boreal forests occur primarily on sand dunes, in glacial lakeplains, and on thin soil over bedrock and cobble of both alkaline and acidic rock types. Farther inland, moderately to poorly drained lakeplain and outwash deposits occasionally support these forests. Within lakeplain, boreal forest is often found in areas with poorly expressed dune and swale topography. Along shorelines, boreal forest often shares an abrupt boundary with coastal communities such as sand and gravel beach, limestone cobble shore, volcanic cobble shore, sandstone cobble shore, limestone bedrock lakeshore, volcanic bedrock lakeshore, sandstone bedrock lakeshore, granite bedrock lakeshore, Great Lakes marsh, open dunes, and Great Lakes barrens. Farther inland boreal forest gradually grades to mesic northern forest, dry-mesic northern forest, or less frequently, rich conifer swamp, granite bedrock glade, limestone bedrock glade, volcanic bedrock glade, or alvar. Coastal boreal forests on the mainland often form narrow, linear bands along the shoreline while archipelagic boreal forests more frequently occupy broader areas of variable shape along island shorelines, especially along the southwestern portions of islands. Nearshore boreal forests occupy peninsulas, former embayments, and coves. Topography of boreal forests ranges widely from gently sloping on lakeplain systems to steep topography on high dune fields, especially where aeolian features have been deposited on moraines.
Soils
Sand, loamy sand, and sandy loam soils are typically moderately acid to neutral, but heavier soils (e.g., silty loam and clay loams) and more acid and alkaline conditions are also found. Boreal forests that occur over bedrock or cobble are often characterized by shallow organic soils or mor humus. Where conifers dominate the canopy, the litter layer is typically more acidic than the underlying organic and mineral soils. Water-retaining capacity of the soils is variable with sandy soils typically being well-drained and soils with heavier texture, such as loams, ranging from moderately drained to poorly drained. Inland boreal forest systems usually occur on moderately to poorly drained lakeplain or outwash.
Natural Processes
Proximity to the Great Lakes results in the moderation of the microclimate of boreal forests with higher humidity, greater snowfall, lower summer temperatures, warmer winter temperatures, and greater summer fog and mist compared to the adjacent inland areas. Natural disturbance regimes are characterized by frequent windthrow, less frequent insect epidemics, and infrequent catastrophic fire. Because boreal forest frequently occurs near the Great Lakes shoreline and thin soils over rock prevent trees from rooting deeply, windthrow and snap-off rates are high; balsam fir is especially susceptible to windthrow and breakage. Spruce budworm (Choristoneura fumiferana) defoliates both spruce (Picea glauca and P. mariana) and balsam fir (Abies balsamea) but tends to be more detrimental to the latter. Interactions of blowdowns, insects, and climate (i.e., droughts) influence fire regimes of boreal forests. Infrequent catastrophic fires are an important disturbance factor, especially in inland boreal forests. Estimations for fire return interval for Canadian and Minnesotan boreal forests range from 50 to 150 years. Given the prevailing landscape position of most Great Lakes boreal forests (e.g., along the shoreline), the fire return interval for these systems was probably greater than 500 years, with fire cycles slightly shorter for inland sites. Large-scale disturbance events in boreal forests can lead to the development of even-aged stands, while small-scale disturbance factors can lead to uneven-aged systems.
Vegetation
The boreal forest flora is circumboreal in distribution with a high degree of floristic homogeneity from site to site. Most species within boreal forests bloom in early spring or summer. The canopy of boreal forests is characterized by a prevalence of conical-shaped evergreens, which often form a closed canopy. The dense tree coverage often results in a scattered understory and sparse ground cover due to the low levels of light transmitted through the canopy and dense sod formed by the extensive network of the shallowly rooted trees. The canopy is dominated by balsam fir (Abies balsamea), white spruce (Picea glauca), and northern white-cedar (Thuja occidentalis), often with lesser amounts of paper birch (Betula papyrifera) and quaking aspen (Populus tremuloides). Dominance shifts toward birch and aspen following fires, large blowdowns, and/or spruce budworm outbreaks, and back toward conifers in the absence of such disturbances. Northern white-cedar dominance is most prevalent in sand dunes and on thin soils over neutral to alkaline bedrock or glacial deposits, such as in the Straits of Mackinac. White spruce is more prevalent on drier sites while balsam fir and cedar are more common on wetter sites; all three of these conifer species increase in importance with time since fire, especially cedar. Additional canopy associates include white pine (Pinus strobus), balsam poplar (Populus balsamifera), and hemlock (Tsuga canadensis), and less frequently black spruce (Picea mariana), red pine (Pinus resinosa), jack pine (Pinus banksiana), and red maple (Acer rubrum). Inland boreal forests are often characterized by an increased canopy component of white pine, hemlock, and deciduous species. Mountain maple (Acer spicatum), striped maple (A. pensylvanicum), American mountain ash (Sorbus americana), and mountain ash (S. decora) are characteristic of the subcanopy and understory. Where aspen and/or birch dominate the canopy, conifers are prevalent in the subcanopy and understory. Additional understory or tall shrub species include round-leaved dogwood (Cornus rugosa), tag alder (Alnus incana), and soapberry (Shepherdia canadensis). Characteristic low shrubs include American fly honeysuckle (Lonicera canadensis), bearberry (Arctostaphylos uva-ursi), Canadian yew (Taxus canadensis), prickly gooseberry (Ribes cynosbati), Canada blueberry (Vaccinium myrtilloides), bush honeysuckle (Diervilla lonicera), common juniper (Juniperus communis), thimbleberry (Rubus parviflorus), and dwarf raspberry (R. pubescens). Ground flora includes species from both mesic northern forest and northern swamp communities such as red baneberry (Actaea rubra), wild sarsaparilla (Aralia nudicaulis), sedges (Carex deweyana and C. eburnea), bluebead lily (Clintonia borealis), goldthread (Coptis trifolia), bunchberry (Cornus canadensis), woodfern (Dryopteris spp.), large-leaved aster (Eurybia macrophylla), fragrant bedstraw (Galium triflorum), Menzies’ rattlesnake plantain (Goodyera oblongifolia), creeping rattlesnake plantain (G. repens), wintergreen (Gaultheria procumbens), twinflower (Linnaea borealis), Canada mayflower (Maianthemum canadense), false mayflower (M. trifolium), naked miterwort (Mitella nuda), partridge berry (Mitchella repens), northern wood sorrel (Oxalis acetosella), bracken fern (Pteridium aquilinum), gay wings (Polygala paucifolia), rose twisted stalk (Streptopus lanceolatus), starflower (Trientalis borealis), and violets (Viola spp.). Ram’s head lady’s-slipper (Cypripedium arietinum, state special concern) and dwarf lake iris (Iris lacustris, federal/state threatened) are uncommon, but characteristic. Mosses and usnea lichens are often abundant due to favorable, moist conditions. Clubmosses, such as stiff clubmoss (Spinulum annotinum), running ground pine (Lycopodium clavatum), and ground pine (Dendrolycopodium obscurum), are often locally abundant, with ground pine more common following fire. Mosses, liverworts, usnea lichens, and saprophytic fungi often are common due to favorable, moist conditions.
For information about plant species, visit the Michigan Flora website.
Plant Lists
Graminoids
- sedges (Carex deweyana and C. eburnea)
- blue wild-rye (Elymus glaucus)
- false melic (Schizachne purpurascens)
Forbs
- red baneberry (Actaea rubra)
- trail-plant (Adenocaulon bicolor)
- wild sarsaparilla (Aralia nudicaulis)
- calypso (Calypso bulbosa)
- bluebead lily (Clintonia borealis)
- goldthread (Coptis trifolia)
- ram’s head lady-slipper (Cypripedium arietinum)
- large-leaved aster (Eurybia macrophylla)
- fragrant bedstraw (Galium triflorum)
- Menzie’s rattlesnake plantain (Goodyera oblongifolia)
- creeping rattlesnake plantain (Goodyera repens)
- twinflower (Linnaea borealis)
- Canada mayflower (Maianthemum canadense)
- starry false Solomon-seal (Maianthemum stellatum)
- false mayflower (Maianthemum trifolium)
- partridge berry (Mitchella repens)
- naked miterwort (Mitella nuda)
- one-flowered pyrola (Moneses uniflora)
- broad-leaved twayblade (Neottia convallarioides)
- one-sided pyrola (Orthilia secunda)
- northern wood-sorrel (Oxalis acetosella)
- sweet-coltsfoot (Petasites frigidus)
- gay-wings (Polygala paucifolia)
- green shinleaf (Pyrola chlorantha)
- lesser pyrola (Pyrola minor)
- twisted-stalks (Streptopus spp.)
- starflower (Trientalis borealis)
Ferns
- wood ferns (Dryopteris spp.)
- oak fern (Gymnocarpium dryopteris)
- bracken fern (Pteridium aquilinum)
Fern Allies
- ground-pines (Dendrolycopodium dendroideum and D. obscurum)
- shining clubmoss (Huperzia lucidula)
- running ground-pine (Lycopodium clavatum)
- stiff clubmoss (Spinulum annotinum)
Lichens
- usnea lichens (Usnea spp.)
Mosses
- dicranum moss (Dicranum montanum)
- stair step moss (Hylocomium splendens)
- largetooth calcareous moss (Mnium spinulosum)
- big red stem moss (Pleurozium schreberi)
- oncophorus moss (Oncophorus wahlenbergii)
- ostrich-plume moss (Ptilium crista-castrensis)
- shaggy moss (Rhytidiadelphus triquetrus)
Shrubs
- tag alder (Alnus incana)
- mountain alder (Alnus viridis)
- bearberry (Arctostaphylos uva-ursi)
- bunchberry (Cornus canadensis)
- round-leaved dogwood (Cornus rugosa)
- bush honeysuckle (Diervilla lonicera)
- wintergreen (Gaultheria procumbens)
- common juniper (Juniperus communis)
- Canadian fly honeysuckle (Lonicera canadensis)
- prickly gooseberry (Ribes cynosbati)
- skunk currant (Ribes glandulosum)
- thimbleberry (Rubus parviflorus)
- dwarf raspberry (Rubus pubescens)
- soapberry (Shepherdia canadensis)
- snowberry (Symphoricarpos albus)
- Canada yew (Taxus canadensis)
- tall bilberry (Vaccinium membranaceum)
- Canada blueberry (Vaccinium myrtilloides)
- oval-leaved bilberry (Vaccinium ovalifolium)
Trees
- balsam fir (Abies balsamea)
- striped maple (Acer pensylvanicum)
- red maple (Acer rubrum)
- mountain maple (Acer spicatum)
- paper birch (Betula papyrifera)
- white spruce (Picea glauca)
- black spruce (Picea mariana)
- jack pine (Pinus banksiana)
- red pine (Pinus resinosa)
- white pine (Pinus strobus)
- balsam poplar (Populus balsamifera)
- quaking aspen (Populus tremuloides)
- American mountain-ash (Sorbus americana)
- mountain-ash (Sorbus decora)
- northern white-cedar (Thuja occidentalis)
- hemlock (Tsuga canadensis)
Noteworthy Animals
Selective browsing by moose in the Upper Peninsula of Michigan can result in the alteration of species composition, community structure, and ultimately forest successional patterns of boreal forests. On sites with spruce and balsam fir, moose preferentially browse on balsam fir, retarding fir vertical growth, limiting fir abundance, and imparting a competitive advantage to spruce. As a predominantly coastal system, Michigan boreal forest and associated communities provide critical feeding, roosting, and perching habitat for migrating shorebirds, waterfowl, and songbirds in the spring. The majority of shrubs found within boreal forest have fleshy fruit, an important food source for birds such as grosbeak, crossbill, warblers, and white-throated sparrow. Paleontologists believe that mastodons (Mammut americanum, extinct) were associated with spruce-dominated forests and that spruce was a dietary staple.
Rare Plants
- Calypso bulbosa (calypso orchid, state threatened)
- Carex concinna (beauty sedge, state special concern)
- Cypripedium arietinum (ram’s head lady’s-slipper, state special concern)
- Disporum trachycarpum (northern fairy bells, state threatened)
- Iris lacustris (dwarf lake iris, federal/state threatened)
- Luzula parviflora (small-flowered woodrush, state threatened)
- Phacelia franklinii (Franklin’s phacelia, state threatened)
- Piperia unalascensis (Alaska orchid, state special concern)
- Pterospora andromedea (pine-drops, state threatened)
- Viburnum edule (squashberry or mooseberry, state threatened)
- Viola epipsila (northern palustrine violet or marsh violet, state threatened)
Rare Animals
- Accipiter gentilis (northern goshawk, state special concern)
- Alces alces (moose, state threatened)
- Canis lupus (gray wolf, state threatened)
- Falcipennis canadensis (spruce grouse, state special concern)
- Falco columbarius (merlin, state threatened)
- Haliaeetus leucocephalus (bald eagle, state threatened)
- Lynx canadensis (lynx, state endangered)
- Pandion haliaetus (osprey, state threatened)
- Picoides arcticus (black-backed woodpecker, state special concern)
- Polygonia gracilis (hoary comma, state special concern)
- Proserpinus flavofasciata (yellow-banded day-sphinx, state special concern)
- Pseudacris triseriata maculata (boreal chorus frog, state special concern)
- Sistrurus c. catenatus (eastern massasauga, federal candidate species and state special concern)
- Sorex fumeus (smoky shrew, state special concern)
- Vallonia gracilicosta albula (land snail, state special concern)
Biodiversity Management Considerations
As a predominantly coastal system, boreal forest and associated communities provide critical feeding, roosting, and perching habitat for migrating shorebirds, waterfowl, and songbirds in the spring. Numerous rare and unique species are associated with boreal forest. When the primary conservation objective is to maintain biodiversity in boreal forests, the best management is to leave large tracts unharvested and allow natural processes (e.g., windthrow, insect defoliation, and fire) to operate unhindered and stochastically generate a range of successional stages. It is crucial to allow dead and dying wood to remain within these systems to become snags, stumps, and fallen logs. Within areas managed solely for biodiversity, resource practitioners should refrain from salvage harvesting following fire, wind, and insect disturbance. Salvage logging, especially following fire, can severely diminish nutrient pools and site productivity in addition to reducing structural heterogeneity.
Chronically high deer densities over the last half-century (or more) have altered tree recruitment, community structure, and floristic composition of Great Lakes forests. Woody plant species unpalatable to deer or tolerant of browsing (e.g., aspens, balsam fir, and red maple) have increased, while species intolerant of deer browsing have decreased (e.g., cedar, hemlock, white pine, yellow birch, and Canadian yew). Reducing deer browse pressure on cedar is a special concern in the Straits of Mackinac, where cedar serves as a canopy dominant.
Currently, logging, shoreline development, and excessive browsing by deer are the primary threats to boreal forests. Few invasive species are presently established in boreal forest but instead pose a potential future threat. Monitoring and control efforts to detect and remove invasive species before they become well established is critical to the long-term viability of the community. Particularly aggressive invasive species that may threaten boreal forest diversity and structure include garlic mustard (Alliaria petiolata), glossy buckthorn (Frangula alnus), Dame’s rocket (Hesperis matronalis), common buckthorn (Rhamnus cathartica), multiflora rose (Rosa multiflora), autumn olive (Elaeagnus umbellata), Eurasian honeysuckles (Lonicera morrowii, L. tatarica, and L. xbella), multiflora rose (Rosa multiflora), and Norway maple (Acer platanoides).
Variation
Following disturbance, boreal forests are characterized by an increased canopy component of early-successional species such as aspen and paper birch. Inland boreal forests typically have more white pine, hemlock, and deciduous species in the canopy compared to coastal boreal forests. Dominance by northern white-cedar is most prevalent on sand dunes and thin soils over neutral to alkaline bedrock or glacial deposits. Coastal boreal forests occurring along the mainland often form narrow, linear bands while archipelagic boreal forests often occupy broader areas of variable shape along island shorelines.
Similar Natural Communities
Rich conifer swamp, mesic northern forest, hardwood-conifer swamp, dry-mesic northern forest, Great Lakes barrens, limestone bedrock glade, granite bedrock glade, volcanic bedrock glade, and wooded dune and swale complex.
Places to Visit
- Cap's Cabin and Waugoshance Point, Wilderness State Park, Emmet Co.
- Isle Royale National Park, Keweenaw Co.
- Keweenaw Point, Baraga State Forest Management Unit, Keweenaw Co.
- Lighthouse Point Boreal Forest (Bois Blanc Island), Gaylord State Forest Management Unit, Mackinac Co.
- Point Betsie, The Nature Conservancy (Zetterberg Preserve at Point Betsie), Benzie Co.
Relevant Literature
- Bergeron, Y., S. Gauthier, V. Kafka, P. Lefort, and D. Lesieur. 2001. Natural fire frequency for the eastern Canadian boreal forest: Consequences for sustainable forestry. Canadian Journal of Forest Research 31: 384-391.
- Cohen, J.G. 2007. Natural community abstract for boreal forest. Michigan Natural Features Inventory, Lansing, MI. 22 pp.
- Darlington, H.T. 1940. Some vegetational aspects of Beaver Island, Lake Michigan. Papers of the Michigan Academy of Science, Arts, and Letters 25: 31-37.
- Flakne, R. 2003. The Holocene vegetation history of Isle Royale National Park, Michigan, USA. Canadian Journal of Forest Research 33: 1144-1166.
- Frelich, L.E., and P.B. Reich. 1995. Spatial patterns and succession in a Minnesota southern boreal forest. Ecological Monographs 65(3): 325-346.
- Greene, D.F., J.C. Zasada, L. Sirois, D. Kneeshaw, H. Morin, I. Charron, and M.-J. Simard. 1999. A review of the regeneration dynamics of North American boreal forest tree species. Canadian Journal of Forest Research 29: 824-839.
- Harman, J.R., and J. Plough. 1986. Asymmetric distribution of coniferous trees on northern Lake Michigan Islands. East Lakes Geographer 21: 24-33.
- Hix, D., B.V. Barnes, A.N. Lynch, and J.A. Witter. 1987. Relationship between spruce budworm damage and site factors in spruce-fir-dominated ecosystems of western Upper Michigan. Forest Ecology and Management 21: 129-140.
- Johnson, E.A. 1992. Fire and vegetation dynamics: Studies from the North American boreal forest. Cambridge University Press, New York, NY. 129 pp.
- Johnstone, J.F., and F.S. Chapin. 2006. Effects of soil burn severity on post-fire tree recruitment in boreal forest. Ecosystems 9: 14-31.
- Kneeshaw, D., and Y. Bergeron. 1998. Canopy gap characteristics and tree replacement in the southeastern boreal forest. Ecology 79(3): 783-794.
- Kneeshaw, D., and S. Gauthier. 2003. Old growth in the boreal forest: A dynamic perspective at the stand and landscape level. Environmental Review 11(1): 99-114.
- Maycock, P.F., and J.T. Curtis. 1960. The phytosociology of boreal conifers — Hardwood forests of the Great Lakes region. Ecological Monographs 30: 1-35.
- McCarthy, J., and G. Weetman. 2006. Age and size structure of gap dynamic old-growth boreal forest stands in Newfoundland. Silva Fennica 40(2): 209-230.
- McCullough, D.G., R.A. Werner, and D. Neumann. 1998. Fire and insects in northern and boreal forest ecosystems of North America. Annual Review of Entomology 43:107-127.
- Pastor, J., B. Dewey, R.J. Naimain, P. McInnes, and Y. Cohen. 1993. Moose browsing and soil fertility in the boreal forests of Isle Royale National Park. Ecology 74(2): 467-480.
- Risenhoover, K.L., and S.A. Maass. 1987. The influence of moose on composition and structure of Isle Royale forests. Canadian Journal of Forest Research 17: 357-364.
- Rowe, J.S., and G.W. Scotter. 1973. Fire in the boreal forest. Quaternary Research 3: 444-464.
- Rutkowski, D.R., and R. Stottlemyer. 1993. Composition, biomass, and nutrient distribution in mature northern hardwood and boreal forest stands, Michigan. American Midland Naturalist 130(1): 13-30.
For a full list of references used to create this description, please refer to the natural community abstract for Boreal Forest.
More Information
Citation
Cohen, J.G., M.A. Kost, B.S. Slaughter, D.A. Albert, J.M. Lincoln, A.P. Kortenhoven, C.M. Wilton, H.D. Enander, M.E. Anderson, M.R. Parr, T.J. Bassett, and K.M. Korroch. 2025. Michigan Natural Community Classification [web application]. Michigan Natural Features Inventory, Michigan State University Extension, Lansing, MI. Available https://mnfi.anr.msu.edu/communities/classification. (Accessed: July 21, 2026).
Kost, M.A., D.A. Albert, J.G. Cohen, B.S. Slaughter, R.K. Schillo, C.R. Weber, and K.A. Chapman. 2007. Natural Communities of Michigan: Classification and Description. Michigan Natural Features Inventory, Report No. 2007-21, Lansing, MI.